Yaddaş konsepsiyası uzun müddətdir bir çox psixoloq, psixiatr və fizioloqun maraq dairəsidir. Etdiyimiz hər şeyə təsiri olan bu konsepsiya illərdir fərqli cəhətlərdən idarə olunur və işləməsi ilə bağlı müxtəlif modellər irəli sürülür. Texnologiyanın bu qədər irəliləmədiyi dövrlərdə “xəstələr” ən böyük məlumat bələdçisi idilər. Bunlardan ən məşhurlarından biri H.M-dir. H.M, uzun illərdir ağır epilepsiya olan bir xəstədir. 1953-cü ildə həkimlərin tövsiyəsi ilə təcrübə üsulu ilə xəstənin medial temporal lobundan (hipokampusun yerləşdiyi beyin hissəsi) toxuma çıxarıldı. Epilepsiya azalsa da, xəstənin yeni xatirələr yarada bilmədiyi (bu xəstəlik anterograd amneziyası adlanır, xəstə 2-3 dəqiqə əvvəl yaşadığı hadisələri xatırlamır), ancaq uşaqlıq xatirələrini aydın şəkildə xatırlaya biləcəyi müşahidə edildi. Xəstədən 2008-ci ildə ölənə qədər yaddaş haqqında çox şey öyrənildi. Bu günlərdə neyrogörüntüləmə cihazlarının inkişafı ilə yaddaşın hansı fiziologiyaya sahib olduğunu və beynin anatomik olaraq hansı hissələri ilə əlaqələndiyini anlamağa çalışırıq. Bu yazıda yaddaşı qısaca izah etdikdən sonra yaddaşın necə işlədiyini öyrənmək üçün xüsusi olaraq təklif olunmuş iki vacib model üzərində dayanacağıq.
Yaddaş nədir? Yaddaş, müxtəlif anatomik quruluşların (amigdala, hipokampus, prefrontal korteks) bir-biri ilə əlaqəli işlədiyi, görmə, eşitmə və məkan məlumatları kimi müxtəlif növ məlumatları duyğu orqanlarımızla əlaqələndirib istifadə edə biləcəyimiz kompleks bir sistemdir. Yaddaşın üç əsas funksiyası olduğu düşünülür: Kodlama (Encoding): Duyğu orqanları tərəfindən alınan məlumatların saxlanıla bilməsi üçün işlənməsidir. Saxlama (Storage): Məlumatın müəyyən bir müddət ərzində depoda saxlanılmasıdır. Xatırlama ( Retrieval) : istifadə edilə bilməsi üçün saxlama sahəsindən məlumatların alınmasıdır, buna geri çağırılma da deyilir. Bundan əlavə, yaddaşın fərqli xüsusiyyətlərini nəzərə alaraq bir neçə sinifə bölmək olar:
• Saxlama vaxtına görə: qısamüddətli yaddaş (short-term memory), uzunmüddətli yaddaş (long-term memory).
• Məlumatların şüurlu səviyyəyə çata biləcəyinə görə: açıq yaddaş (explickt), gizli yaddaş (implicit)
• Bilik xüsusiyyətlərinə görə: epizodik yaddaş, semantik yaddaş bunlardan bəziləridir.
Bəs, bu kompleks və fərqli sistemlər arasında nə əlaqə var? Hansı şərtlərdə bir-biri ilə əlaqə qururlar? Bu nöqtədə yaddaşın işləməsi ilə əlaqədar irəli sürülən iki mühüm modelə rast gəlirik: Multi Store Model (Modal Model) və İşləyən Yaddaş ( Working Memory) .
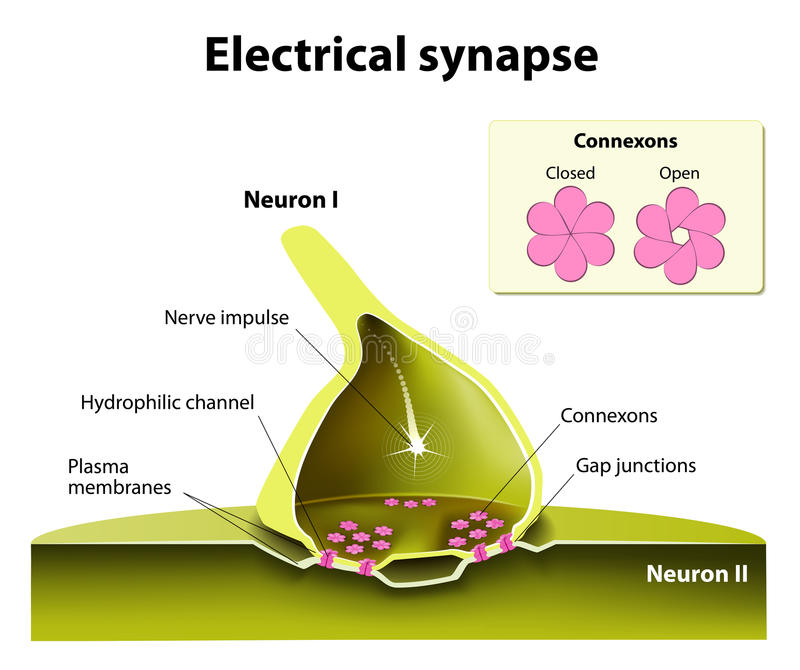
Multi Store Model (Modal Model): 1968-ci ildə Atkinson və Shiffrin tərəfindən təqdim olunan bu model, əslində hamımızın tanıdığımız anlayışlar arasındakı əlaqələri izah edən bir modeldir. Bu modelə görə 3 saxlama sahəsi var. Məlumat bu saxlama sahələrinə ardıcıl olaraq daxil olur və digərinə keçir. Bu dövrü demək olar ki kompüterdəki giriş-çıxış sistemi ilə müqayisə edə bilərik. Məlumat bir saxlama sahəsinə daxil olur və çıxır və digər saxlama sahəsinin girişinə çevrilir. Modeldə nəzərdə tutulan 3 sahənin hamısının xüsusi kodlaşdırma tərzi, saxlama qabiliyyəti və saxlama müddəti var.
Duyğusal Yaddaş (Sensory Memory):
Saxlama sahələrindən birincisi duyğusal yaddaşdır. Bu saxlama zonasında hisslərimizlə aldığımız məlumat, hansı mənada alındığında duyğuya xas bölgədə saxlanılır. Məlumat hər duyğudan əldə olunduğundan fərqli kodlaşdırma formaları (eşitmə, semantik, vizual) burada birlikdə saxlanıla bilər. Bu saxlama sahəsindəki məlumatlar yarım saniyəyə qədər saxlanılır və xüsusi maraq doğurursa, oradakı məlumatlar qısa müddətli yaddaşa köçürülür.
Qısa müddətli yaddaş (Short- term memory):
Qısamüddətli yaddaşda məlumatlar əsasən səs kodlaması ilə kodlanır və məhdud tutuma malikdir (7 +/- 2 obyekt). Məlumat burada yarım dəqiqəyə qədər saxlanıla bilər. Başqa sözlə, bu saxlama sahəsindəki məlumatlar da müvəqqətidir, lakin duyğu yaddaşından fərqli olaraq burada saxlanılan məlumatlar istifadə edilə bilər. Beləliklə şüur səviyyəsində xatırlana bilər. Modelə görə, məlumat yalnız məlumatı təkrarlayaraq uzunmüddətli yaddaşa ötürülə bilər. Ancaq son illərdə aparılan təcrübələrlə bu keçidin o qədər də sadə olmadığını və fərqli parametrlərin də (motivasiya, strategiya) rol oynadığını bilirik.
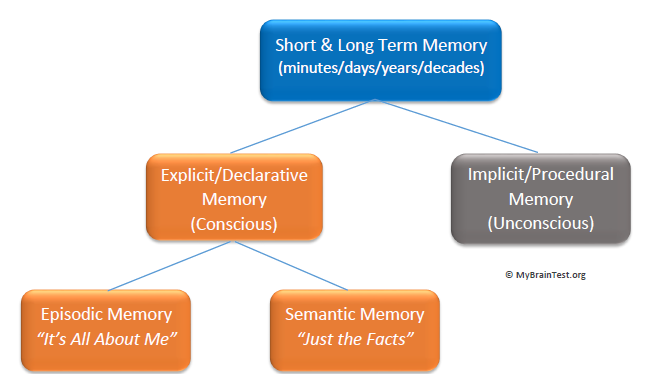
Şəkil 1. Qısa müddətli yaddaş sxemi
Uzun müddətli yaddaş (Long- term memory): Uzunmüddətli yaddaşda məlumat məhdudiyyətsiz bir müddətə saxlanıla bilər. Digər yaddaşlardan fərqli olaraq bu yaddaşın tutumu da məhdud deyil. Model, uzunmüddətli yaddaşın iki alt yaddaş sistemindən ibarət olduğunu söyləyir. Bunlar açıq və gizli yaddaşdır. Açıq yaddaşdakı məlumatlar bilavasitə izah edilə bilər. Və açıq yaddaş daha iki alt vahidə bölünür. Bunlar epizodik və semantik yaddaşdır. Semantik (qavranılan) yaddaş anlayışları, mənaları, qaydaları, məlumatların saxlanılması (“Azərbaycanın paytaxtı Bakıdır” məs.) funksiyasını yerinə yetirir; Epizodik yaddaşda bir vəziyyət və bu vəziyyət haqqında məlumatlar (“10 il əvvəl İU konsertindəydim” kimi) saxlanılır. Örtülü yaddaşda isə şüur səviyyəsinə çatmayan, izah edə bilmədiyimiz və yaxud bildiyimizi bildiyimiz məlumatlar saxlanılır. Buna refleksiv yaddaş da deyilir. Məsələn, üzməyi ilk dəfə öyrənəndə üzmə məlumatları açıq yaddaşdadır. Suyun üstündə durmaq üçün əllərimizi və ayaqlarımızı daim hərəkət eydirməyimiz lazım olduğunu öyrənirik, şüurlu şəkildə tətbiq edirik, amma üzərkən bu bilik şüur səviyyəsinə çatmır, xatırlamırıq, amma var, tətbiq etməyə davam edirik.
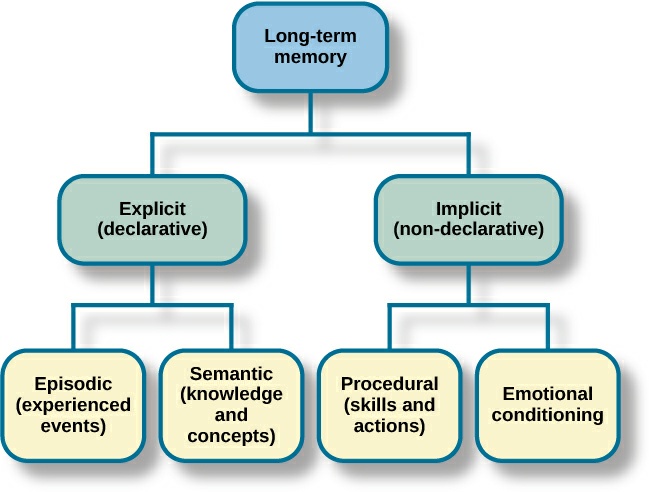
Şəkil 2. Uzunmüddətli yaddaş sxemi
İşləyən yaddaş ( Working Memory) : 1974-cü ildə Baddeley və Hitch tərəfindən yaradılan bu yaddaş modeli bəzən qısamüddətli yaddaşla sinonim olaraq istifadə olunur (qısamüddətli yaddaş məhdud tutuma malikdir və məlumatı müvəqqəti saxlayır), lakin qısa müddətli yaddaşdan böyük fərqlə fərqlənir: burada məlumat sadəcə saxlanılmır həm də işlənə bilir (ing: “məlumatla manipulyasiya”). Bu xüsusiyyətləri ilə idrak prosesslərinin tamamlanmasında və gündəlik həyatımızda böyük rolu olan bir sistemdir. Dörd əsas alt bölmədən ibarətdir:
• Fonoloji döngü
• Vizual və Məkan lövhəsi (Visuospatial Sketchpad)
• Mərkəzi İdarəetmə
• Epizodik bufer
Bu alt hissələrin hər biri fərqli xüsusiyyətlərə malikdir və bir-birləri ilə birbaşa və ya dolayı yolla əlaqə qura bilərlər.
Fonoloji döngü : Bu bölmə 2 alt komponentdən ibarətdir: fonoloji depo və artikulyasiya məşqi (eşitmə məşqi). Fonoloji saxlama, birbaşa eşitmə qabiliyyətində olan məlumatların bir neçə saniyəyə saxlanıldığı məhdud tutumlu (5-8 maddə) bir anbardır. Səs məşqi (bir növ daxili səs) bu depodakı məlumatların bərpa olunmasına, yenilənməsinə və saxlanmasına imkan verən bir müddətdir. Birbaşa səsdən alınan məlumat avtomatik olaraq fonoloji anbarda saxlanıla bilsə də, fərqli xüsusiyyətlərə sahib məlumatların bu anbara keçməsi üçün məlumat səs məşqi ilə eşitmə formasına çevrilməlidir. Məsələn, birisi ilə söhbət edərkən qulaqla aldığımız məlumat birbaşa fonoloji anbarda qeyd edilə bilər, ancaq bir kitab oxuduğumuz zaman eşitmə məşqiylə alınan məlumatları burada saxlaya bilərik. Bu komponent modelin ən çox anlaşılan hissəsidir, komponenti dəstəkləmək üçün bir çox təcrübələr aparılmışdır. Qısaca desək, eşitmə oxşarlığı burada saxlanılan əşyaların sayını artırsa da, semantik oxşarlıq əhəmiyyətli bir təsir göstərmədi (uzunmüddətli yaddaşda semantik loxşarlıq daha çox üstünlük təşkil edir). Söz uzunluğu anında xatırlanan sözlərin sayında azalma təsiri göstərir. Eşitmə söndürmə təcrübələrindən də geniş məlumat əldə edilmişdir. Bu təcrübələrdə iştirakçılardan təcrübədən bir-biri ilə əlaqəsi olmayan bir sözü fasiləsiz və səsləndirmə testində ucadan səsləndirməsi istəndi. Bu şəkildə eşitmə məşqinin funksiyası yatırılır. Təcrübənin sonunda söz uzunluğunun saxlama müddətinə təsiri aradan qaldırıldı. Yadda qalan sözlərin sayı da əhəmiyyətli dərəcədə azaldı, ancaq yox olmadı. Bu, saxlamanın müxtəlif mümkün yollarının olduğunu göstərir.
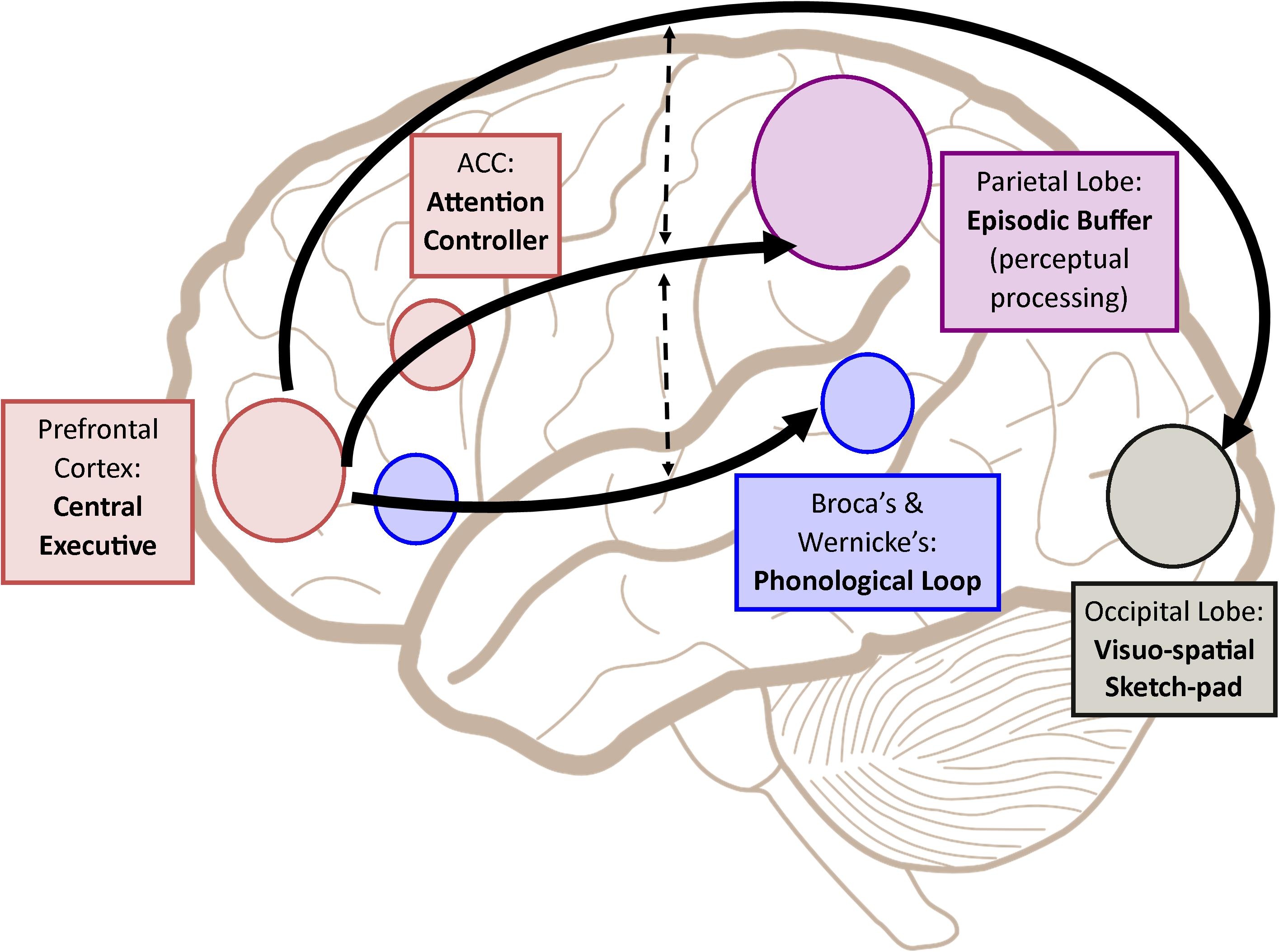
Şəkil 3. İşləyən yaddaş modeli
Vizual və Məkan lövhəsi- Bu bölmədə vizual və məkan (məkan) məlumatları saxlanılır. Təcrübələrdə məkan qavrayışımızı bastıran tapşırıqların vizual iş yaddaşını, görmə qavrayışımızı bastıran tapşırıqların məkan iş yaddaşını təsir etmədiyi müşahidə edilmişdir. Bu saxlama sahəsinin 2 alt bölməyə ayrılması bu təcrübələr tərəfindən dəstəklənir. Başqa sözlə, vizual və məkan məlumatları müstəqil anbarlarda saxlanılır və bu məlumatların işlənməsi də fərqlidir. Vizual iş yaddaşının xarici dünya qavrayışımızda mühüm rolu var. Vizual işləyən yaddaş, gördüyümüz mühitin yalnız müəyyən hissəsini təsirli şəkildə əks etdirə bilər. Bu, ‘dəyişiklik korluğu’ adlanan bir fenomenə səbəb olur. Bu fenomen 1997-ci ildə Rensik tərəfindən belə izah edilmişdir: Uzun müddətli yaddaşda yığılmış vizual mənzərənin məlumatları mərkəzi məlumat sahəsini (mərkəzi maraq bölgələrini) müəyyənləşdirir, bu diqqətimizi yönəldir və məlumatların vizual iş yaddaşına keçməsinə imkan verir. Bu səbəbdən xarici aləmdəki dəyişiklikləri qəbul etmə qabiliyyətimiz böyük ölçüdə bu sahələrlə məhdudlaşır. Diqqət burada mühüm rol oynayır, diqqət etdiyimiz sahədə dəyişiklikləri qəbul edirik. Avtomatik göz hərəkətlərimizin məkan iş yaddaşının səmərəli işləməsində və məkan məlumatlarının saxlanmasında əhəmiyyəti bir çox təcrübə ilə dəstəklənmişdir.
Mərkəzi idarəetmə- işləyən yaddaş modelinin ən vacib hissəsi kimi qəbul olunsa da, modelin ən az anlaşılan və ən az elmi hissəsidir. Bu bölmənin diqqətə nəzarət, konsentrasiya, işləmə və digər komponentlərə məlumat paylamaqla əlaqəli olduğu düşünülür. Digər komponentlərdən fərqli olaraq, tamamilə bir saxlama sahəsi kimi deyil, digər saxlama sahələri (fonoloji döngü, vizual və məkan lövhəsi, uzunmüddətli yaddaş) ilə birlikdə işləyən bir idarəetmə mexanizmi kimi düşünmək olar. Məlumatların işlənməsinin (məlumatların dəyişdirilməsi, digər məlumatlarla inteqrasiya edilməsi) bu vahiddən asılı olduğu təcrübələr tərəfindən dəstəklənmişdir. Bu işləyən yaddaşın qısamüddətli yaddaşdan böyük bir fərqidir. Mərkəzi idarəetmə diqqətimizi yönəldir, eyni zamanda bir çox məlumat arasında seçim etməyə imkan verir və bu məlumatların bəzilərinə üstünlük verir.
Epizodik bufer- fərqli kodlaşdırma növlərinin birlikdə istifadə oluna biləcəyi məhdud tutumlu bir saxlama sahəsidir. Bu saxlama sahəsi eyni zamanda fərqli məlumatların (fərqli hisslərdən əldə edilən, fərqli saxlama sahələrində saxlanılan, fərqli kodlaşdırma növləri ilə kodlanmış məlumatların) və bu inteqrasiya olunmuş məlumatların saxlanılmasında rol oynayır.
Yazar : Yusubova Şəbnəm








Şərh yaza bilmək üçün giriş etməlisiniz.